![]()
La bacteria rebelde
autora: Pascale Cossart
artículo publicado en The Scientist
traducción: Paolo Natale y Miguel Vicente

Moviéndose a través del citoplasma dejando un rastro de actina polimerizada, activando un arsenal de factores de virulencia mediante cambios en la estructura del ARN, o almacenando el código para transcribir el ARN en el lado contrario del ADN, Listeria dicta sus propias normas de supervivencia.
A mediados de los años ochenta, después de estar trabajando varios años en el instituto Pasteur sobre la estructura de proteínas y las interacciones entre ADN y proteína, tenía la oportunidad de cambiar proyecto de investigación y estudiar bacterias patógenas. Junto con mi colaboradora Brigitte Gicquel seleccioné dos sistemas en los que trabajar: la bacteria causante de la tuberculosis, una enfermedad que afecta a alrededor de 9 millones de personas al año, o Listeria, una bacteria que en los Estados Unidos afecta anualmente a unas 2.500 personas y con solo unas 500 muertes al año. Elegí Listeria.
A mí me parecía un organismo modelo perfecto. Al contrario de Mycobacterium tuberculosis, Listeria parecía fácil de manejar, se podía manipular genéticamente, crecía rápido y tenía un ciclo vital interesante. Por entonces se habían hecho muchos estudios sobre patógenos extracelulares y parecía muy interesante estudiar bacterias patógenas intracelulares. Según iba obteniendo más datos, me asombró cómo, una vez que Listeria ha entrado en el interior de una célula, se reviste con una capa de actina, una proteína producida por el hospedador, y parece echarse a volar por el citoplasma como si fuese un supervillano en miniatura. A continuación, en vez de emerger al espacio extracelular por la membrana citoplásmica de la célula del hospedador, como lo hacen otras bacterias, Listeria atraviesa las membranas citoplasmáticas y entra directamente en la célula vecina. Así puede diseminarse directamente a los órganos que ataca: el cerebro y la placenta, sitios que en los mamíferos normalmente están muy protegidos contra la invasión por bacterias. Además, al eludir el espacio extracelular Listeria se hace, al menos en parte, invisible para el radar del sistema inmunitario.
En 1989, Lewis Tilney y Dan Portnoy descubrieron la causa del movimiento a propulsión de Listeria en el interior de la célula. Y este movimiento no lo realiza su flagelo. Este orgánulo, similar a un látigo, es el principal propulsor de Listeria en el ambiente, antes de entrar en el cuerpo del hospedador junto con alimentos contaminados. Pero en el momento en que entra en el cálido intestino del hospedador, la bacteria se desprende de él . Comparando la velocidad de la polimerización de la actina en la célula hospedadora con la distancia recorrida, los investigadores imaginaron que la bacteria avanzaba por la célula construyendo, peldaño a peldaño, una escalera de actina.
La continuación de estos descubrimientos provocó una carera para descubrir el gen que permite a Listeria polimerizar la actina. Pero no era esto lo que a mí me interesaba. Mi laboratorio trabajaba para identificar los genes responsables de la virulencia de la bacteria. Había obtenido una serie de mutantes de Listeria, y con mi colaboradora Edith Gouin, estaba buscando clones que careciesen de la enzima fosfolipasa. Sospechaba que esta enzima interviene en la virulencia ayudando a la bacteria a lisar las membranas que cruza. Una vez encontrado el mutante, nos dimos cuenta enseguida de que la bacteria estaba fuertemente atenuada de otras maneras. Cuando mi postdoc Christine Kocks infectó las células con este mutante, Listeria no pudo formar una cola de actina y desplazarse por el citoplasma. Sin querer participar en la carrera, ¡la habíamos ganado! De hecho la mutación se localizaba en el gen responsable de la movilidad mediante la actina. Ese gen formaba parte de un operon que también contenía el gen de la fosfolipasa, explicando así, por qué ambos genes fueron afectados por la mutación. A este nuevo gen lo llamamos actA(1), gen de Actina A, porque pensé que un proceso tan interesante y peculiar necesitaría la actividad de múltiples genes que encontraríamos más adelante. Es curioso, solo se ha identificado un gen que controla esta característica específica de esta bacteria.
Persiguiendo las respuestas a las preguntas que despertaban mi curiosidad, he dejado que Listeria me conduzca hacia nuevos campos de la Biología, como por ejemplo en los últimos años hacia los ARN no codificantes y la regulación mediada por ARN. Al igual que para moverse en la célula ésta poco convencional bacteria utiliza la actina, Listeria ha desarrollado métodos muy ingeniosos para utilizar los transcritos de ARN que no se traducen.
La bacteria aprovecha todos los componentes del sistema de defensa celular del hospedador en su propio beneficio. Se oculta dentro de las células, secuestra receptores e interfiere con las rutas reguladoras. Listeria es muy adaptable, se encuentra tan feliz lo mismo creciendo en la nevera o en el suelo a 4°C como en el cuerpo a 37°C. En su hospedador se adapta rápidamente al ambiente pobre en oxígeno y se convierte en patógeno. Es esta transición entre la vida saprofita, viviendo de las plantas en descomposición, y la de un patógeno capaz de causar una enfermedad, la que me ha interesado cada vez más.
Desde 1986 hemos estudiado la virulencia utilizando técnicas de biología molecular, bioquímica, y por infección de células en cultivo. También hemos usado ratones que infectábamos por vía intravenosa, pero este modelo era muy artificial, ya que Listeria no infecta a ratones. (Su tropismo se limita a humanos y a animales de granja como vacas y ovejas.) Descubrimos que la versión humana del receptor cadherina E, que es una molécula de adherencia necesaria para mantener la integridad de las uniones entre las células epiteliales, participa en el proceso de entrada de la bacteria. Actua como receptor de una proteína de Listeria llamada internalina A (InlA), obligando a las células del hospedador que producen cadherina E a tragarse la bacteria. Aunque la mayoría de los animales producen cadherina E, la variación de un solo aminoácido entre la cadherina E humana y la de ratón es bastante para que los ratones sean resistentes a la infección de Listeria por vía oral .
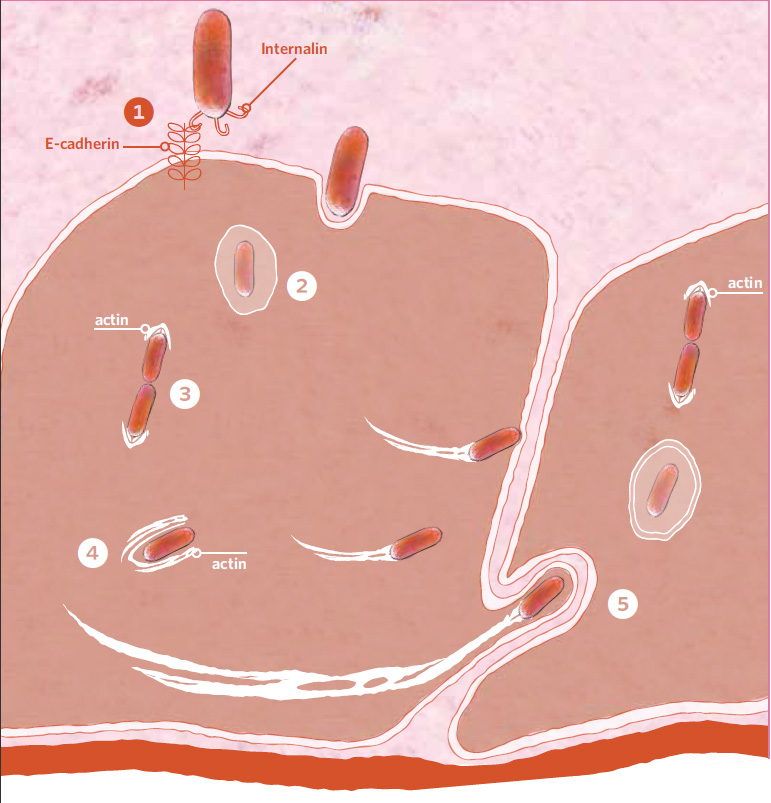
Los trucos que Listeria saca de la chistera
Una vez en el intestino Listeria produce internalina, una molécula de adhesión que interacciona con la cadherina E del hospedador. 1. La bacteria es tragada por la membrana del hospedador 2. Escapándose de la vesícula, Listeria se multiplica 3, cada nueva bacteria nueva queda envuelta en una nube de actina 4. Listeria se lanza por la célula formando una cometa de filamentos de actina y acaba perforando la membrana de una célula adyacente 5. en la que es engullida repitiendo los pasos del 3 al 5.
Para evitar este problema desarrollé, junto con mis colaboradores Marc Lecuit y Charles Babinet, un modelo animal adecuado produciendo la cadherina E humana en el intestino del ratón (2). Fué la primera vez que se usaron ratones humanizados para estudiar una enfermedad bacteriana y que reveló ser un método eficaz. Podemos infectar el ratón por via oral para reproducir la vía de la infección en humanos y así hemos estudiado cómo la bacteria atraviesa la barrera intestinal. Hace poco mejoramos nuestro modelo creando un ratón que solo expresa la cadherina E humana en vez de la de ratón y lo hemos utilizado para estudiar el paso de Listeria por la barrera placentaria.
Según fuimos identificado más factores de virulencia de Listeria notamos que solo se producían a 37°C, la temperatura del cuerpo humano. A temperatura ambiente, no se expresaban ninguno de los genes de virulencia. Aventuramos la hipótesis de que un interruptor, o un factor de transcripción que regula los genes principales de la virulencia, podría estar regulado por temperatura.
Encontramos que en realidad el interruptor se basa en la estructura del ARN mensajero de PrfA (3). A menos de 37°C, la secuencia terminal del ARN se repliega formando una horquilla que hace inaccesible la secuencia de iniciación para que se una el ribosoma y la traduzca. Cuando sube la temperatura, el ARN se despliega y se traduce produciendo el factor de transcripción PrfA, el cual dispara el conjunto de genes de virulencia (véase el gráfico abajo). A este elemento de ARN que actúa al principio de la transcripción le puse, junto con mi postdoc Jörgen Johansson, el nombre de termosensor (3).
Por esta época, alrededor de 2002, los ARNs pequeños emergían como reguladores génicos importantes en eucariotas y procariotas. Pero nuestro termosensor la verdad es que no se comportaba como la mayoría de los ARNs pequeños. Se parecía a lo que se habían llamado elementos ribointerruptores, que forman una horquilla dentro de los ARNs mensajeros inhibiendo su traducción. Hasta nuestro descubrimiento, se creía que todos los ribointerruptores se disparaban por pequeñas moléculas mas que por la temperatura.
¿Qué otros trucos escondía Listeria en su manga para sobrevivir dentro del hospedador humano?
Cuantas más publicaciones leía sobre este asunto, más me intrigaba el poder del ARN. En 2005, ya se había publicado mucho sobre el papel de los ARNs pequeños en los animales y plantas, pero no tanto sobre su función en las bacterias. Mi laboratorio llevó a cabo un proyecto para investigar los ARNs no codificantes de Listeria analizando los transcritos de la bacteria cuando estaba en fase estacionaria (sin alimento), en fase de crecimiento, y cuando está en la sangre como patógeno. Intentamos averiguar qué secuencias se relacionan con la regulación del comportamiento de la bacteria en los principales momentos cruciales de su ciclo de vida y entender cómo cambia de ser una bacteria inofensiva a ser patógena.
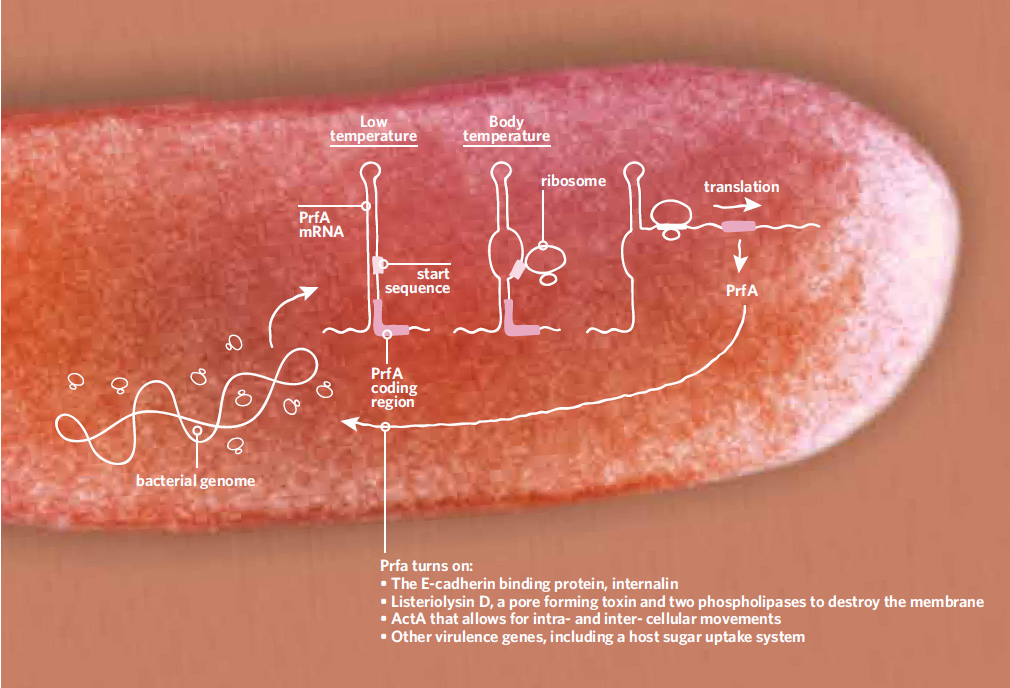
Un termosensor para la virulencia
El gen prfA funciona como interruptor principal de los genes de virulencia de Listeria. A temperaturas bajas (izquierda), cuando la bacteria vive en materia vegetal en decomposición, la secuencia de iniciación del ARN mensajero de PrfA está plegada en forma de horquilla. Cuando la temperatura sube a 37°C, la temperatura del cuerpo (centro), la horquilla se abre y queda accesible la secuencia de iniciación a la que se une el ribosoma iniciando la traducción del mensajero del gen (derecha).
Caracterizamos los transcritos con un “microarray en baldosa”, es un chip de ADN que puede detectar todos los transcritos que la bacteria codifica en cualquiera de las dos cadenas del ADN. Como varios años antes ya habíamos secuenciado y anotado el genoma completo de Listeria, pudimos mapear cada operón del genoma y con cuidado identificar sus secuencias inicial y terminal.
Nuestra búsqueda identificó como mínimo 50 ARNs pequeños que podrían intervenir en la regulación génica, entre ellos dos factores de virulencia (4). Aunque tanto nosotros como otros habíamos ya identificado unos 20 ARNs pequeños que regulan genes de Listeria – y que también aparecieron en esta búsqueda – no se había encontrado ningúno para controlar la virulencia.
Lo que más nos sorprendió es que había varios transcritos largos de ARN que procedían del lado contrario a la cadena que codifica los genes. Si te encuentras en una cadena del ADN un gen que codifica una proteína, nunca miras a la cadena opuesta. Y bien, habíamos identificado estos transcritos antisentido largos pero no sabíamos muy bien para que servían la mayoría de ellos. Una de estas cadenas antisentido contenía una región 5´ no codificante y una región que codifica un regulador transcripcional del gen de la flagelina (5). Este descubrimiento añadía un nivel de complejidad a la ya muy compleja regulación del flagelo de Listeria.
Junto con Johansson, que ahora dirige un grupo de investigación en la universidad de Umeä, demostré que los ribointerruptores pueden actuar sobre genes que les preceden. Además, cuando un ribointerruptor se produce como un transcrito pequeño, en vez de ser degradado, puede actuar en genes lejanos de la cadena de ADN opuesta. De hecho, demostramos que SreA y SreB, dos ribointerruptores, pueden actuar también sobre el transcrito de PrfA uniéndose al inicio de su mensaje. Los transcritos cortos SreA y SreB se producen cuando en la bacteria hay un exceso de S-adenosil, lo que ocurre al crecer en medio rico en nutrientes. Estos transcritos se unen a pfrA e interfieren con su traducción. Todos estos resultados demuestran que prfA – y la virulencia de Listeria en general- están estrictamente controlados por un conjunto de señales ambientales muy sofisticadas, incluyendo la temperatura y la disponibilidad de nutrientes.
El estudio de la Listeria ha sido una mina de oro para mí y para otros que investigan en este tema. Muchas de las preguntas que nos hicimos condujeron al descubrimiento de conceptos y de mecanismos importantes. Además de tener unos ingeniosos trucos para meterse en las células y proliferar en su interior, Listeria también modifica su metabolismo estimulando los genes que le permiten sobrevivir en el hospedador. Puede por ejemplo drenar los azúcares de su hospedador. Además se evade del sistema inmunitario desacetilando su propio peptidoglicano – un componente de la pared celular que normalmente induce la respuesta inflamatoria. También se adueña de la modificación de las histonas y reprograma la célula hopedadora infectada. Recientemente incluso descubrimos proteínas que Listeria envía al núcleo hospedador para reprogramar la célula. Y ahora hay pruebas de que Listeria manipula rutas similares a la ubicuitina para aumentar la infección.
Listeria es una bacteria que ha desarrollado unos mecanismos asombrosamente creativos para la supervivencia en ambientes diversos. Nunca he lamentado mi elección de estudiar a este microscópico malvado.

Pascale Cossart es profesora del Instituto Pasteur e investigadora del programa internacional del Instituto de Medicina Howard Hughes.
REFERENCIAS
1. C. Kocks et al., 1992. “L. monocytogenes-induced actin assembly requires the actA gene product, a surface protein,” Cell, 68:521–531 .
2. M. Lecuit et al., 2001 “A transgenic model for Listeria: role of internalin in crossing the intestinal barrier,” Science, 292:1722-1725 .
3. J. Johansson et al., 2002 “An RNA thermosensor controls expression of virulence genes in Listeria monocytogenes,” Cell, 110:551–561.
4. A. Toledo-Arana et al., 2009 “The Listeria transcriptional landscape from saprophytism to virulence,” Nature, 459:950–956.
5. E. Loh et al., 2009 “A trans-acting riboswitch controls expression of the virulence regulator PrfA in Listeria monocytogenes,” Cell, 139:770–779.
![]()
Foro del día 26 de enero de 2010 en notiweb
Muy interesante la entrada, a pesar de estar en 4º de Biología desconocía muchos detalles acerca de esta bacteria y que me has aclarado en esta entrada del blog.
Estimada profesora Cossart:
Enhorabuena por el post. Es todo un modelo de publicación científica en blog, rigurosa, y con un atractivo hilo narrativo.
Un saludo,
Jaime
Madame Cossart, sin ser del area microbiológica me interesa saber más acerca de la Listeria, para perfeccionar los sistemas de inocuidad alimentaria de las plantas de proceso de alimentos. ¿A que correo electrónico le puedo escribir?.
Gracias, atte.,
Oscar Castro R.
[…] Quien quiera entender mejor el comportamiento de la listeria le recomiendo leer el excelente post publicado por Manuel Vicente la bacteria rebelde. […]
Excelente.
[…] caso diferente nos lo ofrece un reciente estudio de tres cepas de Listeria monocytogenes procedentes de brotes de listeriosis en ganado caprino de la provincia de Jiangsu, al norte de […]