![]()
Plantas micoheterótrofas y micorrizas
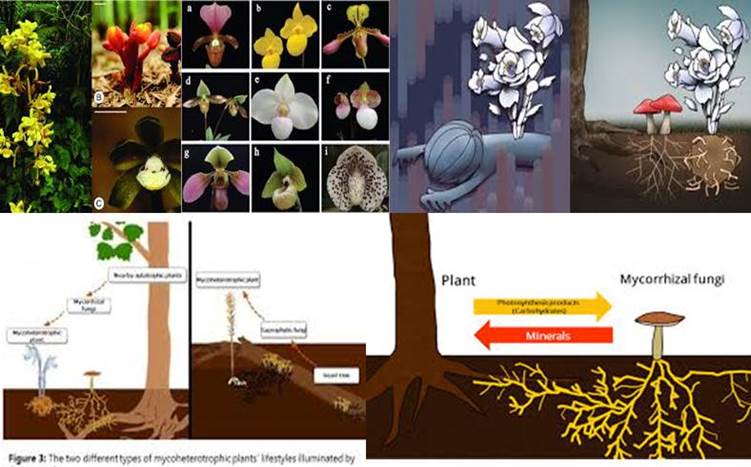
Orquídeas y Micorrizas, pero no suelos: Fuente Imágenes Google
Mis conocimientos de botánica se encuentran ya obsoletos, y mucho más de lo que pensaba. Al seleccionar la nota de prensa sobre micorrizas sobre la que pivota el post de hoy, entendía que hablaríamos de suelos, y en parte es así aunque colateralmente. ¡Craso error! Existen varios tipos de micorrizas al margen de la clásica clasificación de endomicorrizas y ectomicorrizas del medio edáfico, de las que tanto os hemos hablado en este blog. Y es que el término micorriza se aplica a la relación simbiótica entre las raíces de las plantas y ciertos hongos. Sin embargo existen especies epifitas que crecen sobre otros sustratos no edáficos (madera, fragmentos de roca, etc.), que también las atesoran, como esas maravillosas orquídeas y bromelias. Al ir leyendo la noticia he ido topando con términos como plantas heterotróficas, plantas micoheterótrofas, plantas epifitas o epifíticas, hemiepífita primarias, hemiepífitas secundarias, parásitas epirrizas, etc. Empero además en la noticia de abajo se menta: “Las especies micoheterotróficas que explotan hongos ectomicorrícicos deberían tomar fotosinatos recientemente sintetizados, mientras que las plantas micoheterotróficas que dependen de hongos saprotróficos deben obtener carbono de fuentes más antiguas, es decir, como la madera muerta (…) las plantas micoheterotróficas pueden explotar los hongos micorrícicos y saprotróficos (….) Además, a pesar de que el término «micoheterótrofo» ha reemplazado el término mal utilizado «saprófito», ¡algunas plantas micoheterotróficas son indirectamente saprotróficas! . Uffff. Los científicos buscamos definiciones científicas y clasificaciones tan claras y directas como sea posible. Sin embargo, conforme nuestros conocimientos avanzan, el árbol de la vida se va transformando en casi una intrincada red de relaciones difíciles de ordenar e incluso de visualizar gráficamente. Las micorrizas son hongos que amplían la prospección por las plantas (u otros sustratos) en las que se anclan y obtienen su alimento, o parte del mismo. Su red de ramificaciones genera un incremento enorme en la exploración de la superficie de sus hábitats, por lo que no debería haberme extrañado que cuando no sea el suelo, lo sea el substrato vital que conforma su hábitat.
La grandiosa belleza de la vida y la naturaleza no suele permitir que la reduzcamos de forma sencilla, generándose momentos de la historia un tanto complejos. Puede ser frustrante para un científico, pero no deja de asombrarnos y maravillarnos. Un buen día un investigador, o grupo de ellos, logra imponer algo de orden con nuevas indagaciones, empero tarde o temprano, otros encuentran anomalías en los constructos teóricos de la época y volvemos a empezar a intentar simplificar la nueva maraña, en la medida de lo posible, por cuanto tanta complejidad…..
Me asombra también que sigamos hablando de darwinismo a secas, cuando la evolución de la vida constata, una y otra vez esas, que esas asociaciones entre unas formas de vida y otras es un motor tan potente como las desdichadas frases del “pez grande se come al chico” o “sólo sobreviven los más aptos”. “La unión hace la fuerza” tiene tanto peso como la descarnada perspectiva ofrecida por Darwin. Es decir el lamarckismo y el darwinismo son dos fuerzas, del tipo del Yin y del Yang, o dicho de otro modo, las dos caras de una mismo moneda, léase aquí la vida. Pero sobre estos temas ya hablamos sobradamente en nuestra bitácora hace mucho tiempo.
Os dejo con el material que he recopilado para que vosotros mismos os hagáis vuestra composición de lugar sobre estos temas. Hoy tengo la mente un tanto confusa e igual reitero otro “craso error”.
Juan José Ibáñez
Continúa…..
Some non-photosynthetic orchids consist of dead wood
by Staff Writers; Kobe, Japan (SPX) Jan 24, 2020
Botanists have long held a fascination for heterotrophic plants, not only because they contradict the notion that autotrophy (photosynthesis) is synonymous with plants, but also because such plants are typically rare and ephemeral. However, it is still a matter of debate as to how these plants obtain nutrition.
A research team consisting of Kobe University’s Associate Professor SUETSUGU Kenji (of the Graduate School of Science’s Department of Biology), Research Fellow MATSUBAYASHI Jun (of Japan Society for the Promotion of Science) and Professor TAYASU Ichiro (of the Research Institute for Humanity and Nature), has investigated the carbon age (the time since the carbon was fixed from atmospheric CO2 by photosynthesis) in some non-photosynthetic mycoheterotrophic plants. Many orchids have lost photosynthetic ability and evolved an enigmatic mycoheterotrophic lifecycle.
Mycoheterotrophic plants usually obtain carbon from other photosynthetic plants through a shared mycorrhizal fungal network, while some mycoheterotrophs are believed to obtain carbon from decaying litter or dead wood by parasitizing saprotrophic fungi. However, traditional approaches have provided only indirect evidence of such nutrient transportation from dead organic matter to plants.
The current study examined the utility of radiocarbon measurements to distinguish the fungal exploitation pattern of mycoheterotrophs. Mycoheterotrophic species exploiting ectomycorrhizal fungi should take up recently synthesized photosynthates, while mycoheterotrophic plants dependent on saprotrophic fungi must obtain carbon from older sources, i.e., dead wood.
Therefore, the research team calculated the carbon age of mycoheterotrophic plants, using the radiocarbon emitted from atmospheric nuclear bomb tests carried out in the 1950s and 1960s as a tracer.
Through this methodology, they revealed that the carbon in some mycoheterotrophic orchids dated from over ten years prior to the sampling period. This indicates that these orchids are relying on the 14C-enriched bomb carbon from dead wood via saprotrophic fungi. Therefore, they concluded that mycoheterotrophic plants can exploit both mycorrhizal and saprotrophic fungi, which are essential components for terrestrial ecosystems.
In addition, even though the term «mycoheterotroph» has replaced the formerly misapplied term «saprophyte,» some mycoheterotrophic plants are indirectly saprotrophic! The finding overturns the traditional view and opens a new perspective for understanding how these intriguing plants have become ecologically and evolutionarily successful.
Algunas orquídeas no fotosintéticas consisten en madera muerta
Por los escritores del personal; Kobe, Japón (SPX) 24 de enero de 2020
Los botánicos han mantenido durante mucho tiempo una fascinación por las plantas heterotróficas, no solo porque contradicen la noción de que la autotrofia (fotosíntesis) es sinónimo de plantas, sino también porque estas plantas son típicamente raras y efímeras. Sin embargo, todavía sigue siendo tema de debate cómo estas plantas obtienen nutrimentos necesarios para crecer y sobrevivir.
Un equipo de investigación formado por el profesor asociado SUETSUGU Kenji de la Universidad de Kobe (del Departamento de Biología de la Escuela de Graduados de Ciencias), el investigador MATSUBAYASHI Jun (de la Sociedad Japonesa para la Promoción de la Ciencia) y el profesor TAYASU Ichiro (del Instituto de Investigación para la Humanidad y la Naturaleza), ha investigado la edad del carbono (el tiempo desde que el carbono fue fijado a partir del CO2 atmosférico por fotosíntesis) en algunas plantas micoheterotróficas no fotosintéticas. Muchas orquídeas han perdido la capacidad fotosintética y han desarrollado un ciclo de vida micoheterotrófico enigmático.
Las plantas micoheterotróficas generalmente obtienen carbono de otras plantas fotosintéticas a través de una red de hongos micorrícicos compartidos, mientras que se cree que algunos micoheterótrofos obtienen carbono de la materia orgánica en descomposición o de la madera muerta parasitando hongos saprotróficos. Sin embargo, los enfoques tradicionales solo han proporcionado evidencia indirecta de dicho transporte de nutrientes desde la materia orgánica muerta a las plantas.
El estudio actual examinó la utilidad de las mediciones de radiocarbono para distinguir el patrón de explotación fúngica de los micoheterótrofos. Las especies micoheterotróficas que explotan hongos ectomicorrícicos deberían tomar fotosinatos recientemente sintetizados, mientras que las plantas micoheterotróficas que dependen de hongos saprotróficos deben obtener carbono de fuentes más antiguas, es decir, madera muerta.
Por lo tanto, el equipo de investigación calculó la edad de carbono de las plantas micoheterotróficas, utilizando el radiocarbono emitido por las pruebas de bombas nucleares atmosféricas llevadas a cabo en los años 50 y 60 como marcador.
A través de esta metodología, revelaron que el carbono en algunas orquídeas micoheterotróficas databa de más de diez años antes del período de muestreo. Esto indica que estas orquídeas dependen del carbono de la bomba enriquecida con 14C de la madera muerta a través de hongos saprotróficos. Por lo tanto, concluyeron que las plantas micoheterotróficas pueden explotar los hongos micorrícicos y saprotróficos, que son componentes esenciales para los ecosistemas terrestres.
Además, a pesar de que el término «micoheterótrofo» ha reemplazado el término mal utilizado «saprófito», ¡algunas plantas micoheterotróficas son indirectamente saprotróficas! El hallazgo anula la visión tradicional y abre una nueva perspectiva para comprender cómo estas intrigantes plantas se han vuelto ecológica y evolutivamente exitosas.
Research Background
Mutualism, or mutually beneficial interactions between species, is a ubiquitous phenomenon in all ecological systems, and almost all organisms on Earth are involved in at least one mutualistic partnership. Most terrestrial plants, from bryophytes to angiosperms, form mutualisms (interspecific cooperative interactions) with fungi, whereby the plant provides a carbon source in exchange for essential mineral nutrients. These mutually beneficial plant-fungi associations are called mycorrhizal mutualisms.
However, organisms that were originally engaged in mutualisms can sometimes turn into parasites, obtaining the benefits while delivering none in return. In mycorrhizal mutualisms, non-photosynthetic mycorrhizal plants (i.e. mycoheterotrophic plants) are considered as such cheaters since they cannot provide photosynthates to their fungal partners. Indeed, many mycoheterotrophic plants are known to obtain carbon from other photosynthetic plants through a shared mycorrhizal fungal network.
Therefore, despite their achlorophyllous nature, mycoheterotrophic plants are not directly parasitic on other plants, nor do they directly obtain carbon from rotting plant and animal matter, as was once believed. Nonetheless, it is known that fungi play essential roles in terrestrial ecosystems, notably as saprotrophic fungi, which decompose dead wood and decaying litter. Do some mycoheterotrophic plants dependent on these saprotrophic fungi exist? The current research team set out to illuminate this question, utilizing radiocarbon analysis.
El mutualismo, o interacciones mutuamente beneficiosas entre especies, es un fenómeno omnipresente en todos los sistemas ecológicos, y casi todos los organismos en la Tierra están involucrados en al menos una asociación mutualista. La mayoría de las plantas terrestres, desde las briófitas hasta las angiospermas, forman mutualismos (interacciones cooperativas interespecíficas) con hongos, por lo que la planta proporciona una fuente de carbono a cambio de nutrientes minerales esenciales. Estas asociaciones de plantas y hongos mutuamente beneficiosas se denominan mutualismos micorrícicos.
Sin embargo, los organismos que originalmente estaban involucrados en mutualismos a veces pueden convertirse en parásitos, obteniendo beneficios sin entregar ninguno a cambio. En mutualismos micorrícicos, las plantas micorrícicas no fotosintéticas (es decir, plantas micoheterotróficas) se consideran tramposas, ya que no pueden proporcionar fotosintatos a sus socios fúngicos. De hecho, se sabe que muchas plantas micoheterotróficas obtienen carbono de otras plantas fotosintéticas a través de una red de hongos micorrícicos compartidos.
Por lo tanto, a pesar de su naturaleza aclorofílica, las plantas micoheterotróficas no son directamente parásitas de otras plantas, ni obtienen directamente carbono de la materia podrida de plantas y animales, como se creía alguna vez. Sin embargo, se sabe que los hongos desempeñan papeles esenciales en los ecosistemas terrestres, especialmente como hongos saprotróficos, que descomponen la madera muerta y los residuos orgánicos en descomposición. ¿Existen algunas plantas micoheterotróficas que dependen de estos hongos saprotróficos? El equipo de investigación actual se propuso iluminar esta pregunta, utilizando análisis de radiocarbono.
Research Details
Radiocarbon (14C) levels could be useful for precisely estimating the trophic strategies of the symbionts of mycoheterotrophic plants, by providing a direct estimation of the mean carbon age in the biomass. Atmospheric nuclear bomb testing during the mid-20th century increased the 14C concentration in the atmosphere worldwide, the peak of which was around 1963. Subsequently, the atmospheric 14C concentration gradually decreased after the ban on atmospheric nuclear testing in 1963.
Therefore, given that the 14C content of organic matter synthesized by primary producers is the same as the corresponding 14C content of atmospheric CO2, the carbon age (the time since carbon was fixed from atmospheric CO2 by photosynthesis) can be estimated by measuring the concentration of 14C arising from the bomb tests of the 1950s and 1960s. As previously explained, mycorrhizal fungi receive photosynthesized carbon from plants. In addition, the mycoheterotrophic plants exploiting mycorrhizal fungi also obtain carbon from other nearby plants through mycorrhizal network.
Therefore, the research group hypothesized that the 14C values of mycoheterotrophs exploiting mycorrhizal fungi would resemble the 14C values of atmospheric CO2 for the surrounding autotrophic (photosynthesizing) plants, since this carbon should be very recent. On the other hand, the carbon of mycoheterotrophs exploiting saprotrophic fungi (particularly wood-decaying ones) should be older and therefore contain a higher concentration of the 14C that was generated by nuclear tests.
Detalles de investigación
Los niveles de radiocarbono (14C) podrían ser útiles para estimar con precisión las estrategias tróficas de los simbiontes de las plantas micoheterotróficas, al proporcionar una estimación directa de la edad media de carbono en la biomasa. Las pruebas de bombas nucleares atmosféricas a mediados del siglo XX aumentaron la concentración de 14C en la atmósfera en todo el mundo, cuyo pico fue alrededor de 1963. Posteriormente, la concentración atmosférica de 14C disminuyó gradualmente después de la prohibición de las pruebas nucleares atmosféricas en 1963.
Por lo tanto, dado que el contenido de 14C de la materia orgánica sintetizada por los productores primarios es el mismo que el contenido de 14C correspondiente de CO2 atmosférico, la edad del carbono (el tiempo desde que el carbono se fijó del CO2 atmosférico por fotosíntesis) puede estimarse midiendo la concentración de 14C derivado de las pruebas de bombas de los años 1950 y 1960. Como se explicó anteriormente, los hongos micorrícicos reciben carbono fotosintetizado de las plantas. Además, las plantas micoheterotróficas que explotan hongos micorrícicos también obtienen carbono de otras plantas cercanas a través de la red micorrízica.
Por lo tanto, el grupo de investigación planteó la hipótesis de que los valores de 14C de los micoheterótrofos que explotan hongos micorrícicos se asemejarían a los valores de 14C del CO2 atmosférico para las plantas autotróficas (fotosíntesis) circundantes, ya que este carbono debería ser muy reciente. Por otro lado, el carbono de los micoheterótrofos que explotan los hongos saprotróficos (particularmente los que se descomponen en la madera) debería ser más antiguo y, por lo tanto, contener una mayor concentración de 14C que se generó mediante pruebas nucleares.
To investigate these hypotheses, the 14C concentrations of 10 species of mycoheterotrophic plants collected across 10 sites in Japan were measured. Of these ten species, six species (two ericaceous species Monotropastrum humile and Pyrola subaphylla and four orchidaceous species Cephalanthera subaphylla, Chamaegastrodia shikokiana, Neottia nidus-avis and Lecanorchis nigricans) showed low 14C concentrations which were similar to the results for autotrophic plants- confirming that they utilize very recent carbon.
On the other hand, it was revealed that the other four species, which were all types of orchid (Gastrodia elata, Cyrtosia septentrionalis, Yoania japonica and Eulophia zollingeri), contained very high concentrations of 14C, dating from over ten years prior to the sampling period. These research results indicate that some mycoheterotrophs have acquired 14C-enriched bomb carbon from dead wood via saprotrophic fungi. This indicates that some mycoheterotrophic plants do not obtain their carbon by tapping into existing mycorrhizal networks, but recruit saprotrophic fungi into novel mycorrhizal symbioses.
Conclusion
This research illuminated that mycoheterotrophic plants can exploit both mycorrhizal and saprotrophic fungi, which are essential components for terrestrial ecosystems. Many botanists rejected the use of the term ‘saprophyte’ as incorrect and called the plants mycoheterotrophic plants to reflect their unique nutritional dependence on fungal carbon.
In fact, there are no saprophytes that directly feed on dead organic matter. However, the radiocarbon approach provides conclusive evidence that some mycoheterotrophic orchids are indirectly saprophytic and are dependent on wood debris in the forest carbon cycle.
The results of this research will be published online in New Phytologist on January 24, 2020.
Con vistas a investigar estas hipótesis, se midieron las concentraciones de 14C de 10 especies de plantas micoheterotróficas recolectadas en 10 sitios en Japón. De estas diez especies, seis especies (dos especies de ericáceas Monotropastrum humile y Pyrola subaphylla y cuatro especies de orquídeas Cephalanthera subaphylla, Chamaegastrodia shikokiana, Neottia nidus-avis y Lecanorchis nigricans) mostraron bajas concentraciones de 14C que fueron similares a los resultados para plantas autotróficas que confirmaron Utilizan carbono muy reciente.
Por otro lado, se reveló que las otras cuatro especies, que eran todos los tipos de orquídeas (Gastrodia elata, Cyrtosia septentrionalis, Yoania japonica y Eulophia zollingeri), contenían concentraciones muy altas de 14C, que datan de más de diez años antes del periodo de muestreo. Los resultados de estas investigaciones indican que algunos micoheterótrofos han adquirido carbono de bomba enriquecido con 14C de la madera muerta a través de hongos saprotróficos. Esto indica que algunas plantas micoheterotróficas no obtienen su carbono aprovechando las redes micorrícicas existentes, sino que reclutan hongos saprotróficos en nuevas simbiosis micorrícicas.
Conclusión
Esta investigación iluminó que las plantas micoheterotróficas pueden explotar los hongos micorrícicos y saprotróficos, que son componentes esenciales para los ecosistemas terrestres. Muchos botánicos rechazaron el uso del término ‘saprófito’ como incorrecto y llamaron a las plantas micoheterotróficas para reflejar su dependencia nutricional única del carbono fúngico.
De hecho, no hay saprófitos que se alimentan directamente de materia orgánica muerta. Sin embargo, el enfoque de radiocarbono proporciona evidencia concluyente de que algunas orquídeas micoheterotróficas son indirectamente saprófitas y dependen de los residuos leñosos del ciclo del carbono forestal.
Los resultados de esta investigación se publicarán en línea en New Phytologist el 24 de enero de 2020.
Resumen del Artículo original en Español-castellano
Las plantas micoheterotróficas dependen completamente de asociaciones fúngicas para nutrientes orgánicos. Si bien la mayoría de las plantas micoheterotróficas están asociadas con los socios micorrícicos de las plantas verdes circundantes, se cree que algunos micoheterótrofos obtienen carbono de la materia orgánica en descomposición o de la madera muerta al parasitar hongos saprotróficos, basados en experimentos de cultivo y firmas isotópicas 13C y 15N.
La edad del carbono (el tiempo desde que el carbono se fijó a partir del CO2 atmosférico mediante la fotosíntesis) se puede estimar midiendo la concentración de 14C que surge de las pruebas de bombas de los años cincuenta y sesenta. Dado que los hongos micorrícicos obtienen fotosinato de sus socios vegetales y los hongos saprótrofos que descomponen la madera obtienen carbono de fuentes más antiguas, el radiocarbono podría representar una herramienta nueva y poderosa para investigar las fuentes de carbono de las plantas micoheterotróficas.
Demostramos que los valores de Δ14C de los micoheterótrofos que explotan los hongos ectomicorrícicos eran cercanos a 0 ‰, similares a los de las plantas autotróficas. Por el contrario, los valores de Δ14C de los micoheterótrofos que explotan hongos saprotróficos oscilaron entre 110.7 ‰ y 324.8 ‰, debido al carbono de la bomba enriquecida con 14C de la madera muerta a través de hongos saprotróficos.
Nuestro estudio proporciona evidencia que respalda que algunas orquídeas micoheterotróficas dependen de los restos leñosos del bosque. Nuestro estudio también indica que el radiocarbono podría usarse para predecir las estrategias tróficas de los simbiontes fúngicos asociados a micoheterótrofos.
Plantas micoheterótrofas
Sabemos que ciertos hongos necesitan de un huésped para subsistir, en un proceso más o menos conocido denominado micorrización… Como en cualquier relación simbiótica, ambos seres reciben algo a cambio del otro, generalmente la planta proporciona hidratos de carbono y vitaminas que el hongo no puede obtener por sí mismo, y la planta recibe humedad y minerales fruto de la descomposición de la que el hongo es capaz.
Hasta aquí bien, el hecho curioso es que a veces esa relación simbiótica deja de ser útil para uno de los miembros, donde el otro pasa a ser un parásito que no aporta nada a cambio. Es el caso de las plantas micoheterótrofas, un tipo muy curioso de plantas que parasitan a los hongos. La característica más curiosa es que no poseen clorofila, por lo que sus colores suelen ser exóticos. Por otro lado, sus hojas son vestigiales, ya que no tienen ninguna función.
Micoheterotrofia (Wikipedia)
La micoheterotrofia (del griego mico, hongo; heterotrofa, diferente y trofo, que se alimenta) es una relación biológica que se establece entre ciertas clases de plantas y hongos en la que las plantas obtienen, total o parcialmente, sus nutrientes mediante parasitismo sobre el hongo antes que por fotosíntesis. Un micoheterótrofo es la planta parásita en esta relación. La micoheterotrofia está considerada como una relación de explotación, y a los micoheterótrofos se les conoce de un modo informal como los «explotadores de micorrizas». También se refiere a esta relación como micotrofia, aunque este término también se utiliza para las plantas que tienen relaciones micorrizales mutualistas.1
La micoheterotrofia total (u obligada) existe cuando una planta que no realiza la fotosíntesis (una planta que carece de clorofila o lo que es lo mismo carente de una fotosíntesis funcional) obtiene todos sus nutrientes de los hongos que parasita. La micoheterotrofia parcial (o facultativa) existe cuando una planta es capaz de realizar la fotosíntesis, pero parasita a un hongo como aporte suplementario de nutrientes. Las plantas verdes que tienen esta capacidad dual de absorber nutrientes de un hongo por micoheterotrofia y, al mismo tiempo, retienen la capacidad autotrófica que les brinda la fotosíntesis, se denominan mixótrofos.23 También hay plantas, tales como algunas especies de orquídeas, que no fotosintetizan y son micoheterotróficas obligadas durante una parte de su ciclo de vida mientras que fotosintetizan y son micoheterotróficas facultativas o autotróficas para el resto de su ciclo vital.4 Es importante tener en cuenta que otras plantas que no realizan la fotosíntesis y son también aclorofilas (es decir, no presentan clorofila), tales como las especies de los géneros Cuscuta u Orobanche no son micoheterotróficas ya que son plantas parásitas que parasitan directamente el tejido vascular de otras plantas.5
Hasta hace pocos años se creía, de un modo erróneo, que las plantas que no realizan la fotosíntesis conseguían sus nutrientes directamente de la materia orgánica de una manera saprofítica, similar a la que utilizan los hongos. Tales plantas, por lo tanto, fueron llamadas «saprófitas». Ahora se sabe que estas plantas no son capaces de la absorción y digestión directa de la materia orgánica y que para conseguir el alimento deben recurrir al parasitismo micoheterotrófico o directo de otras plantas.67
a interfaz entre la planta y sus socios fúngicos en esta asociación está entre las raíces de la planta y el micelio del hongo. La micoheterotrofia por lo tanto se asemeja mucho a las micorrizas y, de hecho, actualmente se considera que puede haberse desarrollado a partir de micorrizas,6 excepto que en la micoheterotrofia, el flujo de nutrientes se verifica desde el hongo hacia la planta y no al revés.89
Los micoheterótrofos, por tanto, se pueden considerar como epiparásitos, puesto que toman la energía de los hongos los cuales a su vez consiguen su energía de otras plantas vasculares.67 De hecho, muchos micoheterótrofos se encuentran comúnmente en un entramado micorrizal, en el que las plantas utilizan las micorrizas del hongo para el intercambio de dióxido de carbono y nutrientes.7 En estos sistemas, los micoheterótrofos juegan el papel de «explotadores micorrizales», tomando los nutrientes de las micorrizas comunes, pero sin dar nada en contrapartida.6
Las plantas micoheterotróficas, entonces, obtienen la totalidad del carbono desde un hongo, el cual -a su vez- lo obtiene desde una planta verde, fotosintética, con la que vive asociado en una relación mutualística. Se ha demostrado recientemente que en muchas plantas mixotróficas de la familia de las ericáceas una considerable proporción del carbono proviene de los hongos con los que conviven. Así, los arbustos ericáceos de los bosques boreales pertenecientes a la tribu Pyroleae, filogenéticamente relacionados con micoheterótrofos obligados dentro de la subfamilia Monotropoideae, Orthilia secunda, Pyrola chlorantha, Pyrola rotundifolia y Chimaphila umbellata adquieren entre un 10,3 y un 67,5% de su carbono desde los hongos que parasitan, conjuntamente con una parte sustancial del nitrógeno.1011