![]()
Fotocromo, fotomodulación, fotorreceptores y regulación armónica del crecimiento vegetal. (Salvador González Carcedo).
Muchas personas creen que en la fotosíntesis reside únicamente el crecimiento vegetal, mas o menos apoyado por las hormonas y alguna cosilla mas. Decimos que “existe un complejo sistema de captación de radiaciones (fotosistemas) y además, una fase oscura cuya responsabilidad es la de orientar al metabolismo de los vegetales a sintetizar compuestos orgánicos”. Pero una cosa es disponer de compuestos fotosintetizados en la fase oscura de la fotosíntesis, y otra muy distinta es hacer crecer armónicamente a un vegetal y decidir cuando y cómo lo hace.
Como creo que la cosa es complicada para los profanos, vamos a adentrarnos en el mundo de los fotorreceptores lentamente.
Para empezar, decir que existen otros conjuntos moleculares diferentes a las clorofilas que usan radiaciones distintas. Son los fotomodulades o fototransmisores que conforman el Fotocromo. Y podemos definirle como un amplio conjunto de pigmentos sensores (que está aumentando rápidamente), que controlan muy diferentes procesos de desarrollo de la planta, e informan a las plantas de los cambios operados en su entorno ambiental, natural o forzado), optimizando su crecimiento.
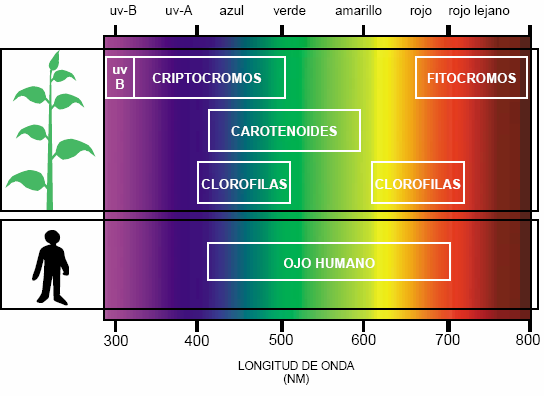
Con el fotocromo se perciben los distintos segmentos del espectro de radiaciones que alcanzan a la planta, evaluables en distintos parámetros como dirección espacial, intensidad, duración y periodicidad. Y efectos concretos derivados del tipo de día (claro, luminoso, nublado, frío, cálido…) que “sufren”, de la estación en la que se encuentra y/o de la cercanía de otras plantas. Como consecuencia de la información recibida, las plantas generan respuestas de adaptación, modulando sus propios procesos, desde la germinación hasta la desetiolización, pasando por el fototropismo o la floración, el crecimiento y la fructificación.
En cuanto a las consecuencias que generan la actividad del Fotocromo, el conjunto de respuestas que afectan al desarrollo y aspecto de la planta, en función de la luz recibe el nombre de “fotomorfogénesis”, pero los conocimientos endógenas que condicionan este proceso abarcan aspectos muy variados (bioquímicos, metabólicos y fisiológicos y hormonales) y su manejo adecuado, tiene grandes consecuencias técnicas, económicas agronómicas y ecológicas, pues no se puede olvidar que la luz es una gran fuente de información para las plantas.
A diferencia de los sistemas fotosintéticos (Cloroplastos) la localización del fotocromo: es ubicua en el reino vegetal, y solo dudosa (su existencia) en los hongos. En algas verdes se cita su localización en el plasmalema, siendo mucho más difícil el estudio de la distribución del fotocromo en plantas superiores. Por técnicas espectroscópicas e inmunocitoquímicas se determinó la ubicación de algunos fitocromos, a nivel tisular y celular. En general, la mayor concentración parece estar en tejidos meristemáticos o en curso temprano de diferenciación tanto en raíces como en tallos, aunque también se lo encuentra en pequeña cantidad en las hojas. En las células se lo encontró asociado al retículo endoplásmico, las mitocondrias e incluso a los etioplastos. En coleóptilos de maíz el fitocromo parece estar unido a membranas.
Haciendo un poco de historia les recordaré que el detección de los primeros fitocromos se realizó hacia 1950 evaluando la acción de la luz sobre el proceso de germinación de semillas de lechuga. El estudio del efecto producido por destellos de luz sobre la germinación de ciertas plantas permitió concluir que mientras la luz roja producía activación de germinación, si se usaban destellos de rojo lejano se inhibía este proceso. El proceso revertía y el último tipo de luz que recibían las plantas era el determinante de la respuesta final.
Veamos los resultados
| Tratamiento lumínico | Germinación |
| Oscuridad | 21% |
| 5 min. R | 92% |
| 5 min. Rojo+10 min. Rojo Lejano | 38% |
| Rojo-Rojo Lejano-Rojo | 88% |
| Rojo-Rojo Lejano-Rojo-Rojo Lejano | 42% |
| 10 min. Rojo Lejano | 40% |
Con estos datos, se determinó que mientras que una luz de 780 nm (Rojo Lejano) tiene efecto negativo una luz de 680 nm (Rojo) ejerce un efecto positivo.
Hoy se sabe que existe un receptor molecular con dos opciones posibles e interconvertibles, de manera que pueden cambiar y revertir de una en otra, según la aplicación de determinadas longitudes de onda. En otras palabras una forma del receptor será activa a efectos fisiológicos y la otra forma será inactiva y en todo caso los datos demuestran que puede alcanzarse un equilibrio de fotoconversión.
A la forma inactiva del fitocromo se denominó Pr mientras que la forma activa es Pfr, siendo esta última la que controla indirectamente múltiples procesos como floración, germinación o movimiento foliar. Además, en las respuestas mediadas por el fitocromo va a tener importancia no sólo el efecto de cada forma por separado sino que la responsabilidad del efecto fisiológico recae sobre la relación Pr/Pfr a esta relación se la conoce hoy como Estado Fotoestacionario.
Para complicar mas las cosas, el conocimiento nos va dando una de la importancia que tienen cada distintos grupo de moléculas sensibles a las radiaciones y un criterio de ubicación de las mismas dentro de los procesos generales de captura de luz.
Así, implicadas en la fotosíntesis (generación de ATP y potencial reductor) aparecen los conjuntos clorofílicos confomados por mezclas de clorofilas a, b. c, d, (capaces de absorber entre los 535 y los 730 n.m.) aliados con los carotenoides, que absorben luz verde y amarilla (400-600 nm), evitando la desestabilización de los PS o fotosistemas de una excesiva agitación molecular (estress lumínico). Ciertamente, la luz afecta en la fotosíntesis a muchos enzimas, de la misma forma que a los sistemas de transferencia de iones y al ritmo de trabajo de enzimas tan importantes como la conocida Rubisco o la menos conocida anhidrasa carbónica (una de las enzimas más veloz), pero no compliquemos la información.
Sin embargo, en el control de la fotomorfogénesis participan otros fotorreceptores que captan y transmiten señales de diferentes regiones del espectro: el receptor de luz ultravioleta-B (280- 320 nm), los criptocromos, que captan la radiaciones del UV-A (320-390 nm) y azul (400-500 nm); y los fitocromos, que perciben la luz roja y roja lejana (700-800 nm). Y siguen apareciendo nuevas moléculas.
Así, en el mundo vegetal, a nivel de organización molecular se han identificado al monos tres grandes clases:
(A) Fitocromos (Phy, bPhy según sean de plantas o bacterias): son conjuntos protéicos complejos, cuyo grupo prostético es un cromóforo (un tetrapirrol lineal), sensibles a la energía que portan las radiaciones entre los 600 y 750 nm, es decir, entre el rojo y el IR (infrarrojo) cercano. Esta sensibilización ocurre gracias a una isomerización “cis <<<=>>> trans” de alguno de sus dobles enlaces.
Existen tres grupos de fotocromos: A (los mas abundantes en plantas etioladas y cuya presencia disminuye en la medida en que se hacen mas verdes) y B y C que aparecen en pequeña cantidad en plantas etioladas y verdes.
Para los mas interesados, recordarles que este fotorreceptor:
· son proteínas diméricas compuestas por dos polipéptidos de 124 KDa.
· cada uno de los monómeros se une de forma covalente a otra molécula (un pigmento llamado fitocromobilina mediante un enlace tioéter.
· en los vegetales, la apo-proteína de los fitocromos contiene dos histidín quinasas llamadas dominios (HKRD1 y HKRD2) en el área del carboxilo terminal y otros dos dominios llamados Per-Amt-Sim (PAS) en el seno del HKRD1 en cuyo seno muestran su función como un dominio de interacción proteína–proteína respondiendo como pequeños módulos ligando.
· Los fitocromos presentan dos sensibilidades distintas a las radiaciones, derivadas de dos estados, cuyo nombre, comportamiento y estabilidad son diferentes: Así la forma Pr: tiene un pico de absorción a 660 nm (color azul) La forma PfR muestra ese pico de absorción a 730 nm (color azul verdoso) (la Pfr se transforma espontáneamente en Pr gracias a la iluminación, con radiaciones del IR cercano, y a la inversa).
· Pf se aloja en el citoplasma, mientras que la forma activa (Pfr) se localiza preferentemente en el núcleo celular.
· La bioquímica nos muestra dos tipos de Pfr distintos, según de que vegetal se trate. El tipo I (forma Pfr fotolábil) y el tipo II (forma Pfr fotoestable). En las plantas etioladas predomina en un 90% el tipo I (fotolábil) En las plantas verdes los tipo I y II aparecen en cantidades iguales. En Arabidopsis se han encontrado 5 fitocromos (PhyA-E) con este cromóforo, pero con sensibilidad diferente a la luz. Así, el PhyA corresponde al tipo I, mientras que el resto (PIB, PhyC, PhyD y PhyE) corresponden al tipo II.
En cuanto a sus acciones sobre las plantas podemos decir que presentan tres tipos de respuesta en el vegetal:
a) Fotorespuestas de nivel bioquímico/metabólico/fisiológico
a) Estimulación de la síntesis de enzimas y proteínas en plantas etioladas e iluminadas, tales como proteínas estructurales, enzimas mitocondriales (glutámico deshidrogenasa), enzimas citoplasmáticas (glucosa 6-fosfato-deshidrogenasa).
b) Síntesis de antocianinas y otros flavonoides.
c) Biosíntesis de giberelinas.
d) Regulación de la síntesis de alcaloides
e) Sincronización de ritmos circadianos.
b) Respuestas fotoperiódicas que afectan a la morfogénesis del vegetal:
a) Iniciación e interrupción de la latencia invernal del brote de las peremnes leñosas.
b) Regulación de la germinación de algunos tipos de semillas.
c) Formación de bulbos en ciertas plantas.
d) Apertura estomal/ crecimiento longitudinal de los estomas
c) Respuestas morfogenéticas no fotoperiódicas
a) Desenrrollamiento de hojas en monocotiledóneas.
b) Inhibición del alargamiento del tallo.
c) Alargamiento de rizoides en musgos.
d) Alteración del geotropismo y fototropismo.
De los fitocromos bacterianos: también podemos decir que participan en la fototaxis de bacterias fotosintetizadoras y de microalgas. También en la expresión genética (e.g. síntesis de pigmentos, síntesis del aparato fotosintético, sincronización del reloj biológico de los ritmos circadianos. etc.,.).
(B) Criptocromos (Cry) son cromoproteinas cuyo grupo cromóforo es una flavina (FAD) unida a una pterina, que les hace sensibles a radiaciones en dos picos en los rangos del UV (350-366 nm) y del azul 450 nm. Se ubican sobre el aminoácido amino terminal de la proteína del dominio relacionado con la fotoliasa fotosintética (dominio PHR), y su área carboxílico terminal conforma un motivo DQXVP-ácido-STATES (DAS).
Sus actividades, de forma paralela a los fitocromos, se centran en
· Regular la síntesis de enzimas y antocianinas
· Favorecer la apertura de estomas.
· Favorecer el crecimiento de tallos.
· Regular el fototropismo.
· Favorecen la diferenciación de los plástidos y su especialización
(C) Fototropinas (Nph1, Phot) son conjuntos moleculares cuya grupo cromóforo es una flavina del tipo FMN que les hace sensibles a radiaciones entre 320-500 nm cuando forma aductos con una molécula de cisteína. Es decir entre la luz azul y la UVA (ultravioleta. Las apo-proteínas de la fototropina contienen dos dominios de unión a cromóforos (LOV1 y LOV2) y otro dominio quinasa en el extremo carboxílico.
En un tipo paralelo de moléculas aparecen las proteínas “BLUF”– (AppA) cuyo grupo cromóforo es otra flavina (FAD) unida a una pterina.
A la hora de interpretar como funciona el conjunto del proceso, hay que delimitar que este tipo de moléculas que configuran el fotocromo desarrollan dos funciones, una sensora y otra reguladora.
Así la función sensora implica la percepción de la señal luminosa incidente y por ello hay que especificar claramente su espectro de absorción y la longitud de onda de máxima absorción a los que trabaja, teniendo en cuenta que estas moléculas conforman complejos orgánicos.
La función reguladora consiste en transferir la información recibida (intensidad y tiempo u orientación, en un contexto general ambiental) a los componentes de la cadena de transducción de la señal, y desde ésta, a los elementos diana, constituidos por otros componentes celulares. Como la luz incidente es policromática, los fitocromos operarán como interruptores de naturaleza molecular, que informan a la planta de la presencia y los cambios en las proporciones relativas de luz (roja y de roja lejana del ambiente, en el caso de los fitocromos), para que acometa las respuestas fisiológicas oportunas.
Pero esto será motivo de otro post.
Saludos cordiales
me gustaria saber mas sobre sus articulos y trabajos
Excelente explicacion sobre el modo de percepcion de luz de la planta, esta super correcto. Soy alumno de la facultad de agronomia de la universidad de Buenos Aires y felicito por este post en este blog. Informacion muy confiable y util.
te felicito por tu explicacion sobre la influencia de la luz en plantas, esta muy concreto y muy facil de entender. soy alumna de la facultad de aronomia en MÉXICO S.L.P. ME GUSTARIA QUE EXPUSIERAS UN TEMA TRATADO CON EL RITMO CIRCADIANO EN PLANTAS.
Dos pregunatas:
1. Existe este término Fotocromo con «o»?
2. La función sensora y reguladora, es del Fotocromo o del Fitocromo.
gracias
Paola